Cell surface specialization

Cell surface is not only divided into the basal, apical and lateral parts within the specialization, which mainly refers to epithelial cells. However, even non-epithelial cells can have surface specializations (cilia, microvilli, etc.).
Basal surface
This is the pole at the basement membrane. The specialization here is basal labyrinth or they also call it basal annealing. In it, the cell membrane runs into the interior of the cell and forms numerous folds (intussusception). mitochondria are then located between the folds, which in this place provide ATP for the transport of ions against the concentration gradient, which is very energy demanding. In the membrane of the labyrinth there are also integral membrane proteins that form ion pumps. We can find it, for example, in the 'proximal channels of the kidneys.
Apical surface
This is half way to the free surface. The specialized formations here are microvilli, stereocilia and cilia (kinocilia).
Microvilli

They are the smallest of all the mentioned apical formations. They are 0.5–1 μm long, then average about 0.1 μm. These are cytoplasmic protrusions bounded by a membrane. Inside, they contain twenty to thirty actin microfilaments in a longitudinal arrangement. Villin and fimbrin form bundles of filaments.
The connection of the microfilaments to the cell membrane is then ensured by another protein called calmodulin. The continued anchoring of microfilaments within the cell is in the terminal network winding under the microvilli, another fulcrum of anchoring is subsequently mediated by the actin-binding protein 'fodrin', which connects the terminal network to the intermediate filaments.
When a large number of microvilli cover the entire apical surface, we call it 'brush border'. Thanks to this, the surface area increases and thus the absorption. We find it on the surface of enterocytes in the lumen of the intestine and in the cells lining the proximal kidney tubules.

Stereocilia
Of the above-mentioned formations, they reach a medium length, which can be up to 7 μm. These protrusions are immobile, but similar in structure to microvilli, they are formed by parallel arranged microfilaments connected by α-actinin. They are thicker at the base and can subsequently branch out, but their diameter does not change during their course. We find them in the ductus epididymidis epididymis and in the inner ear in the hair cells.
Kinocilia
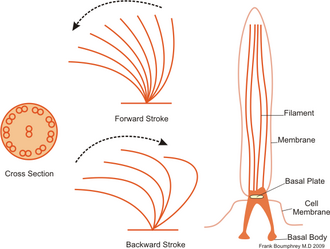
Kinocilia, also called cilia, are mobile cytoplasmic protrusions formed by a complex ciliary apparatus. They are approximately 10 μm long, with an average thickness of about 0.2 μm. It consists of the basal body and the structures connected to it, which are: the transitional part, the free part itself and the terminal part. They can be found (due to their mobility) in the trachea, where they transport exogenous substances from the lungs, and in the fallopian tube, in which they help the movement of the egg into the womb.

- Basal body has the same structure as centriole. It is connected to it by striated roots pointing into the cytoplasm of the cell and also by a striated basal leg running vertically.
- Transitional part of the cilium is the area between the basal body and the free part of the cilium. We do not find central microtubules here, only the ends of microtubules C triplets of the wall of the basal body. Here, they are attached to the membrane, which helps anchoring.

- Own free cilia is already the cytoplasmic protrusion itself covered by a cilia membrane. The center is formed by an axoneme composed of two central microtubules surrounded by a central sheath. Surrounding the central pair are nine double peripheral microtubules. Peripheral microtubules arranged close to each other are continuations of microtubules A and B triplets of the wall of the basal body. Fiber molecules of the tektin protein are stored longitudinally between them. Microtubule A is composed of thirteen protofilaments, microtubule B of eleven protofilaments, which is due to the fact that part of the wall is shared with microtubule A. On microtubule A there are two (inner and outer) 50 nm long arms of dynein, a motor protein with a high ATP activity. The outer arms are more frequent, they are attached to every third tubulin heterodimer, the inner arms are up to every fourth heterodimer. The connection of peripheral microtubules with the central sheath is provided by radial connectors, which are connected to every fourth tubulin heterodimer. Connection of double peripheral microtubules to each other is ensured by nexin. It is 30 nm long but can stretch up to 250 nm.
Cross-section of the cilia, their movement and the overall structure of the cilia.

- The terminal part of the cilia is a short section where double peripheral microtubules change to single ones and gradually disappear. In the same way, the diameter of the eyelash also narrows. We can sometimes find brush-like structures on the surface of the plasmalemma.
Movement of the cilia takes place in one plane, where a rapid bending is followed by a gradual straightening. Movement is ensured by the mutual displacement of adjacent pairs of microtubules, which is created by a temporary connection between the dynein arms of microtubule A of one pair and microtubule B of the other pair. The connections are only short-lived, rapid formation is followed by rapid disconnection - this repetitive event can be compared to the climbing of individual microtubules. The motor of movement is MAPs dynein, whose structure is similar to cytoplasmic MAP1C dynein. One side of MAPs is bound to the A microtubule, the other is made up of three globular particles that have high ATPase activity. ATP binds to them, which releases energy through hydrolysis. The energy changes the configuration of dynein, causing the protein to move and with it the double peripheral microtubules. The movement only takes place in one half of the axoneme, so the resulting movement is performed in only one plane.
Links
Related articles
References
- JUNQUIERA, L. Carlos – CARNEIRO, José – KELLEY, Robert O.. Basics of histology. 1. edition. H & H 1997, 1997. 502 pp. ISBN 80-85787-37-7.
- LÜLLMANN-RAUCH, Renate. Histology. 1. edition. Grada, 2012. 576 pp. ISBN 978-80-247-3729-4.
